

Археология об основании Рима: Новые раскопки проясняют и такой острый дискуссионный вопрос, как дата самого возникновения Рима...

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Археология об основании Рима: Новые раскопки проясняют и такой острый дискуссионный вопрос, как дата самого возникновения Рима...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Основы обеспечения единства измерений: Обеспечение единства измерений - деятельность метрологических служб, направленная на достижение...
Интересное:
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Финансовый рынок и его значение в управлении денежными потоками на современном этапе: любому предприятию для расширения производства и увеличения прибыли нужны...
Наиболее распространенные виды рака: Раковая опухоль — это самостоятельное новообразование, которое может возникнуть и от повышенного давления...
Дисциплины:
|
из
5.00
|
Заказать работу |
Содержание книги
Поиск на нашем сайте
|
|
|
|
Использование эволюционного подхода при изучении различных этапов развития центральной нервной системы позвоночных животных в значительной мере расширяет наши представления об интегративной деятельности мозга, о механизмах поведения и психических функций. В настоящее время сравнительная нейрология, вооруженная электрофизиологическими, морфологическими, авторадиографическими, гистохимическими методами, успешно решает вопросы о функциональном назначении мозговых структур, о межцентральных связях, об афферентном снабжении различных корковых формаций и т. д. Успешность этих исследований во многом определяется правильным выбором объекта экспериментальной работы. Следует отметить, что современные представители всех классов позвоночных формируют условный филогенетический ряд, так как животные, составляющие непрерывную линию эволюции, вымерли. В то же время современные позвоночные большей частью подверглись значительной адаптивной специализации. В связи с этим особого внимания заслуживают те таксоны, которые подверглись меньшей специализации и отражают действительную историю формирования мозга, включающую спинальную, ихтиопсидную, зауропсидную и маммальную стадии развития. Специфические особенности этих стадий описаны в данной главе.
Спинальный этап развития
В ходе эволюции трубчатая система позвоночных животных имеет несколько критических этапов развития, на протяжении которых возникают новые уровни интеграции в ростральных, филогенетически более молодых отделах мозга. Исходным считается спинальный тип центральной нервной системы, представленный у наиболее древних хордовых – бесчерепных. В составе нервной трубки современного бесчерепного – ланцетника, нет дифференцированного головного мозга. Спинной мозг имеет форму длинного тяжа, состоящего из 64 однородных сегментов, от которых отходят задние (чувствительные) и передние (двигательные) корешки. Они выходят из спинного мозга отдельными стволами.
На поперечном срезе спинной мозг ланцетника имеет трапециевидную форму. В центре трапеции проходит спинномозговой канал. Внутреннее строение нервной трубки отличается примитивностью. Четкого разделения на белое и серое вещество не отмечается, однако нервные клетки имеют тенденцию концентрироваться вокруг центрального канала. Белое вещество спинного мозга ланцетника состоит из безмякотных волокон, в основном малого диаметра. Вместе с тем в ростральных (6-11-й сегмент) и каудальных (39-61-й сегмент) отделах нервной трубки располагаются гигантские клетки Родэ (рис. 11), толстые аксоны которых после дугообразного изгиба следуют на значительное расстояние в пределах спинного мозга, формируя проводящие пути, имеющие координационное значение.
К клеткам Родэ в составе задних корешков подходят афферентные аксоны от светочувствительных глазков Гессе, которые локализованы вдоль всей нервной трубки, но преимущественно в передней и хвостовой частях тела ланцетника. Каждый глазок Гессе., состоящий из пигментной и ганглиозной клетки и представляющий собой примитивный фоторецептор, реагирует на световые раздражители, свободно распространяющиеся через прозрачные покровы тела. Возбуждение от фоторецепторов через синаптические переключения достигает клеток Родэ и далее адресуется мотонейронам, аксоны которых иннервируют мышечные пластинки ланцетника. Таким образом, происходит замыкание рефлекторной дуги, лежащей в основе поведенческой реакции – отрицательного фототаксиса.
Сенсорный аппарат ланцетника представлен помимо глазков Гессе реснитчатыми клетками эпителия, которые реагируют на химические, термические и механические раздражители, в частности на вибрационные колебания водной среды. Афференты от этих рецепторных образований в составе задних корешков проникают в спинной мозг ланцетника.
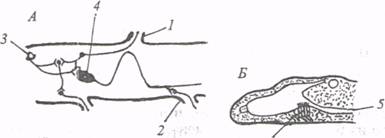
Рис. 11. Расположение нервных элементов в нервной, трубке ланцетника (по: А. И. Карамян, 1976; А.Л.Поленов, 1971):
А – срединная часть и Б– головной конец нервной трубки; 1– задний корешок, 2 – передний корешок, 3 – глазок Гессе, 4 – клетка Родэ, 5 – спинномозговой канал, 6 – инфундибулярный орган
В головной части нервной трубки расположена обонятельная ямка, выстланная клетками чувствительного эпителия. Эти клетки снабжены ресничками и, по мнению ряда авторов, обладают хеморецепторной функцией. Однако вопрос о функциях обонятельной ямки остается мало освещенным. В вентральной стенке переднего конца нервной трубки локализован «инфундибулярный орган», который состоит из крупных клеток, обладающих нейросекреторной функцией. Апикальная часть этих клеток достигает расширенной полости спинномозгового канала, базальный же полюс переходит в отросток, идущий к вентральной поверхности нервной трубки (рис. 11). Цитоплазма клеток инфундибулярного органа заполнена мелкими (100-300 нм) гомориположительными секреторными гранулами. Согласно представлениям А.Л. Поленова (1971), описанная выше структура является гомологом нейросекреторных клеток преоптического или супраоптического ядра гипоталамуса других, более высокоорганизованных позвоночных животных.
На основании структурной специфичности головного отдела нервной трубки ланцетника некоторые авторы (Ф.В. Овсянников, В.Я. Данилевский) считали эту область зачатком головного мозга – архенцефалоном, регулирующим всю рефлекторную деятельность бесчерепных:. Однако в результате физиологических исследований, где анализировали восприятие и простые формы обучения у ланцетника, установлено, что нет достаточных оснований говорить о специфике головного отдела нервной трубки. Так, например, по данным Б.Ф. Сергеева, перерезка нервной трубки в ее центральной части ни в коей мере не отражается на условнорефлекторных реакциях ланцетника. Они вырабатываются одинаково успешно как при раздражении рецепторов головного отдела, так и при стимуляции хвостовой части тела.
Условнорефлекторные реакции формировались при использовании одномодальных раздражителей, например слабого света, создающего освещенность 30 люкс, как условного раздражителя и сильного света (120 люкс) как безусловного подкрепления, После 28-36 сочетаний условного и безусловного раздражителей слабый свет приобретал сигнальное значение и вызывал двигательную реакцию - отрицательного фототаксиса. Вместе с тем при сочетании разномодальных раздражителей (света и электрического тока) условнорефлекторная реакция не вырабатывалась, что свидетельствовало об отсутствии феномена замыкания между нервными элементами, воспринимающими условный и безусловный раздражители. Эти факты говорят о том, что приобретенные реакции ланцетника по своему механизму отличаются от классических условных рефлексов высших позвоночных.
Формирование условных реакций у ланцетника протекает по типу повышения возбудимости афферентных и эфферентных систем спинного мозга при повторном раздражении. В данном случае суммирование следов возбуждения с наличным возбуждением приводит к повышению чувствительности (сенсибилизации) ЦНС, которая начинает реагировать на условные раздражители, бывшие до того подпороговыми. Такие индивидуально приобретенные реакции относятся к категории суммационных рефлексов. У ланцетника суммационные рефлексы характеризуются малой стабильностью и сохраняются не более десяти часов. Из всех видов внутреннего торможения у этих животных вырабатывается только угасательное.
Таким образом, бесчерепным доступны наиболее простые формы обучения, которое осуществляется по механизму суммационного рефлекса. Эти функции реализуются теми отделами нервной трубки, которые имеют сенсорный вход от рецепторов (глазков Гессе) и моторный выход к сократительному аппарату. Надсегментарных (мозговых) систем интеграции в ЦНС бесчерепных нет, и в целом можно говорить о равнозначности всех отделов нервной трубки в плане восприятия и простых форм обучения. Отсутствие выраженной структурной и функциональной специализации характерно для наиболее древнего, спинального этапа развития ЦНС позвоночных животных, согласно предложенному А.И. Карамяном принципу этапности в историческом развитии ЦНС.
|
|
|

Историки об Елизавете Петровне: Елизавета попала между двумя встречными культурными течениями, воспитывалась среди новых европейских веяний и преданий...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...

Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...
© cyberpediasu.com 2017-2026 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!

