Убедительно показано, что у всех физиологических групп хемоавтотрофов ассимиляция СО2 происходит через реакции цикла Кальвина. Выращенные в хемоавтотрофных условиях клетки характеризуются высоким содержанием двух специфичных для данного метаболического пути ферментов - рибулозодифосфаткарбоксилазы и фосфорибулокиназы. Однако у факультативно автотрофных тиобацилл и у водородных бактерий синтез этих двух ферментов часто в той или иной степени подавляется, если клетки выращиваются на органических субстратах. Многие облигатно автотрофные тиобациллы и нитрифицирующие бактерии имеют особые характерные для прокариот образования, карбоксисомы, содержащие рибулозодифосфаткарбоксилазу.
Для осуществления ассимиляции СО2 хемоавтотрофы должны получать и АТФ, и восстановитель (восстановленный пиридиннуклеотид) путем окислительной диссимиляции неорганического субстрата. По урожаю биомассы (таблица 7) можно оценить относительную эффективность их получения разными организмами.
ТАБЛИЦА 7.- Урожай биомассы некоторых хемоавтотрофных бактерий 1
| Организм
| Биомасса
|
| Pseudomonas facilis
Thiobacillus neapolitanus
Thiobacillus ferrooxidans
| 12 г на 1 г-моль Н2
4 г на 1 г-моль S2O32-
0,35 г на 1 г-атом Fe2+
|
1 Урожай выражен в граммах сухой биомассы клеток, синтезированной на грамм-молекулу или грамм-атом окисленного субстрата.
ОКИСЛЕНИЕ МОЛЕКУЛЯРНОГО ВОДОРОДА
В принципе генерация как АТФ, так и восстановленного пиридиннуклеотида не представляет особых сложностей, если окисление неорганического субстрата сопряжено, подобно окислению органических субстратов, с восстановлением НАД+. Часть образованного таким образом НАД.Н может окисляться в дыхательной цепи переноса электронов, что сопровождается синтезом АТФ при окислительном фосфорилировании, а часть использоваться для осуществления восстановительных реакций ассимиляции углерода. По-видимому, именно такая ситуация и имеет место у водородных бактерий, поскольку показано, что некоторые из них обладают растворимой гидрогеназой, которая катализирует реакцию

Этот фермент, выделенный из клеток Nocardia opaca, обладает двумя интересными свойствами. Во-первых, он чувствителен к кислороду и инактивируется на воздухе в нейтральной среде, а во-вторых, для его работы необходимы ионы Ni2+, которые могут быть заменены на Mg2+ (правда, с частичной потерей активности фермента).
Эти особенности гидрогеназы проливают свет на некоторые казавшиеся ранее странными особенности питания водородных бактерий. Так, было известно, что хотя окисляющие водород псевдомонады при выращивании на органических субстратах ведут себя как обычные аэробные организмы, некоторые из них при хемоавтотрофном росте становятся микроаэрофильными и развиваются за счет Н2 только тогда, когда парциальное давление кислорода оказывается значительно ниже, чем при обычных условиях в атмосфере.
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ И СИНТЕЗ ВОССТАНОВЛЕННЫХ ПИРИДИННУКЛЕОТИДОВ У ДРУГИХ ХЕМОАВТОТРОФОВ
Механизмы, посредством которых хемоавтотрофы, отличные от водородных бактерий, удовлетворяют свои синтетические потребности при окислении субстрата, представляются гораздо более сложными. В качестве примера можно рассмотреть бактерии, которые окисляют нитрит. У бактерий этой группы при окислении субстрата высвобождается только одна пара электронов:

Однако потенциал пары NO2- /NO3- (Е'0 =+0,42 В) гораздо выше потенциала, необходимого для осуществления сопряженной реакции
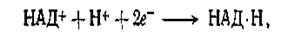
для которой Е'0 составляет —0,32 В.
Полученные из бесклеточных экстрактов Nitrobacter фракции частиц обладают системой переноса электронов и могут катализировать окисление нитрита кислородом. Исследование такой системы показало, что в ходе этой реакции происходит перенос электронов от нитрита на О2 через цитохромы а1 и а3, который сопровождается синтезом 1 моля АТФ на 1 моль окисляемого субстрата (рисунок 8). Таким образом, механизм генерации АТФ при таком типе хемоавтотрофии становится вполне понятным, но способ образования восстановленного пиридиннуклеотида по-прежнему неясен. Проведенные недавно исследования показали, что НАД+ в бесклеточной системе восстанавливается нитритом при добавлении в систему АТФ, который затрачивается на обратный перенос электронов от восстановленного цитохрома а1 (первого акцептора электронов в цепи переноса). Как схематически показано на рисунке 9, для осуществления реакций, приводящих к восстановлению НАД+, требуется несколько молейАТФ. Таким образом, часть энергии, полученной при окислении нитрита молекулярным кислородом (через посредство конечных компонентов цепи переноса), должна быть затрачена на то, чтобы перенести электроны от нитрита против термодинамического градиента и восстановить пиридиннуклеотид.
Полученные недавно данные показывают, что аналогичные процессы обратного переноса электронов требуются и для восстановления пиридиннуклеотидов за счет других неорганических субстратов, которые могут окислять хемоавтотрофы (Fe2+, NH3 и восстановленные соединения серы). Однако ни одна из этих систем не изучена пока так детально, как система окисления нитрита. Более того, во многом невыясненными остаются этапы реакций и биохимические механизмы, которые функционируют при окислении аммиака до нитрита и при окислении соединений восстановленной серы до сульфата. При окислении аммиака (валентность N равна—3) до нитрита (валентность N равна +3) атом азота теряет шесть электронов.
Рисунок 8.- Сопряжение синтеза АТФ с окислением нитрита у Nitrobacter.
.
Рисунок 9. Возможный путь энергозависимого обратного переноса электронов от нитрита к НАД+ у Nltrobacter
Следовательно, этот процесс протекает с участием по крайней мере двух промежуточных продуктов, в которых атом азота имеет промежуточные валентности. Есть веские данные в пользу того, что первый этап этого процесса катализируется оксигеназой, которая присоединяет к аммиаку полученный от О2 атом кислорода, в результате чего образуется гидроксиламин (NH2OH) (валентность N равна-1). Каковы последующие этапы процесса, пока не известно. Возможно, имеет место следующая цепь реакций:

При окислении H2S до сульфата теряется восемь электронов. Поскольку у многих окисляющих H2S хемолитотрофов наблюдается быстрое внутриклеточное накопление элементарной серы, есть все основания считать, что это вещество является первым стабильным промежуточным продуктом, хотя ферментативный механизм его образования неизвестен. Предпоследним промежуточным продуктом является сульфит. Природа реакций, приводящих к его образованию, остается неясной, хотя предложено несколько возможных вариантов. Механизм окисления
сульфита до сульфата и ферменты, участвующие в этом процессе, твердо установлены. Это единственная связанная с окислением неорганического соединения реакция, которая позволяет запасать энергию на уровне субстратного фосфорилирования, поскольку протекает с образованием аденилированного промежуточного соединения лденилилсульфата (АФС):
Так как с помощью реакции, катализируемой аденилаткиназой, из АДФ может быть образован АТФ:
в результате фосфорилирования АМФ на уровне субстрата образуется АТФ.












